i am inserting trigonometry notes which you can read and download , these are the core concepts of trignometry
it starts from very simple pythogaras theorem to the sum and difference formula
We will read about these in detail with pictures and lots of knowledge
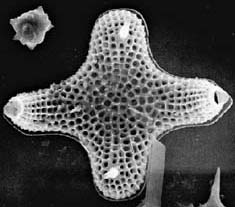
BACILLARIOPHYTA (diatoms)
CHAROPHYTA (stoneworts)
CHLOROPHYTA (green algae)
CHRYSOPHYTA (golden algae)
CYANOBACTERIA (blue-green algae)
DINOPHYTA (dinoflagellates)
PHAEOPHYTA (brown algae)
RHODOPHYTA (red algae)
BACILLARIOPHYTA(DIATOMS)
Bacillariophyta are unicellular organisms that are important components of phytoplankton as primary sources of food for zooplankton in both marine and freshwater habitats. Most diatoms are planktonic, but some are bottom dwellers or grow on other algae or plants.
Except for their male gametes, diatoms lack flagella. Instead many diatoms
achieve locomotion from controlled secretions in response to outside physical and chemical stimuli. Diatoms have unique shells, which serve as their cell wall. The overlapping shells, or frustules that surround the diatom protoplasm are made of polymerized, opaline silica. Identification of diatom species is based on the delicate markings on their frustules, comprising a large number of tiny, intricately-shaped depressions, pores and passageways that bring the diatom’s cell membrane in contact with the environment. Diatom frustules have accumulated over millions of years to form the fine, crumbly substance known as diatomaceous earth, which has a variety of uses (e.g. for filtration and insulation). Diatom remains in both marine and freshwater sediments are also important as indicators of paleo-environmental conditions at the time the sediments were formed.
Bacillariophytes have brownish plastids containing chlorophylls a and c and fucoxanthin. The primary means of reproduction is asexual, by cell division. Most diatoms are autotrophic, but a few are obligate heterotrophs (they must absorb organic carbon) because they lack chlorophyll altogether. Some diatoms even lack their distinctive frustules and live symbiotically in large marine protozoa, providing organic carbon for their hosts.
A diatom. Numbered graduations are 10 micrometres apart
Charophyta are freshwater plants and generally grow anchored to the substratum by rhizoids with a shoot extending upward. The shoot then divides and forms nodes from which a whorl of side filaments projects. Charophyta reproductive structures develop at these nodes and are, along with the biflagellate sperm produced in the male gametangium, quite similar to those of mosses. These similarities have led some scientists to identify the charophytes as ancestors of the mosses. Their green color comes from chlorphylls a and b.
CHLOROPHYTA(GREEN ALGAE)
Most chlorophytes are aquatic, but some green algae can live on the surface of snow, on tree trunks, in soils, or symbiotically with protozoans, hydras or lichen-forming fungi. Numbering about 8,000 species, the chlorophytes range in size from microscopic to quite large. The typical color of plants in the Chlorophyta, resulting from the dominant chlorophyll pigments, is some shade of apple or grass green, although certain species may appear yellow-green or blackish-green due to the presence of carotenoid pigments or high concentrations of chlorophyll. Chlorophytes appear more than a billion years ago in the fossil record.
Calcified green algae, particularly Halimeda spp., are especially important as major contributors of marine sediments. The sparkling white sand beaches of the Caribbean and many other areas in the world are largely the sun-bleached and eroded calcium-carbonate remains of green algae. The deepest occurring, fleshy, erect alga Johnson-sea-linkia profunda, (Littler et al., 1985)was found attached to bedrock at a depth of 157 meters off the Bahamas and is a member of this group.
Green algae have chlorophylls a and b and store starch as a food reserve inside their plastids. Most green algae have firm cell walls composed of cellulose along with other polysaccharides and proteins.
CHRYSOPHYTA (golden algae)
Chrysophytes are photosynthetic, unicellular organisms that are abundant in freshwater and marine environments. Chrysophytes contain chlorophylls a and c, which are masked by the accessory pigment fucoxanthin, a carotenoid. In many ways, golden algae are, biochemically and structurally similar to brown algae. Both golden algae and brown algae store food outside of the chloroplast in the form of polysaccharide laminarin, or chrysolaminarin. In both groups, motile cells have unequal flagella of similar structure.
CYANOBACTERIA (blue-green algae)
Even though the Cyanobacteria are classified as bacteria (lacking a membrane-bounded nucleus) they are photosynthetic and are included among our algal collections. Cyanobacteria played a decisive role in elevating the level of free oxygen in the atmosphere of the early Earth. Cyanobacteria can change remarkably in appearance, depending on the environmental conditions. Blue-green algae are common in soil, in both salt and fresh water, and can grow over a wide range of temperatures. They have been found to form mats in Antarctic lakes under several meters of ice and are responsible for the beautiful colors of the hot springs at Yellowstone and elsewhere. Cyanobacteria can also occur as symbionts of protozoans, diatoms and lichen-forming fungi, and vascular plants. Some blue-greens can fix nitrogen as well as photosynthesize, allowing them to grow with only light, water, a few minerals, and the nitrogen and carbon dioxide in the atmosphere.
Cyanobacteria are different in many important ways from other photosynthetic prokaryotes. Instead of the bacteriochlorophylls found in purple and green bacteria, blue-greens contain chlorophyll a, as in eucaryotic phototrophs, and, produce free oxygen as a byproduct of photosynthesis. Cyanobacteria, however, lack the organized chloroplasts of eukaryotes and have their photosynthetic apparatus distributed peripherally in the cytoplasm.
The variety of striking colors exhibited by Cyanobacteria are a result of their major light-gathering pigments, the phycobilins, that are bound to protein granules, (phycobilisomes), that are attached to the photosynthetic membranes.
Large blooms of freshwater Cyanobacteria may produce toxins that can kill livestock. Other forms (Spirulina) are grown commercially and marketed as a high-protein dietary supplement.
PYRROPHYTA/DINOPHYTA (dinoflagellates)
[Dinoflagellata - Pyrrophyta - Pyrrhophyta] * (Current names in use by various authorities)
The division Pyrrophyta (from the Greek "pyrrhos" meaning flame-colored) comprises a large number of unusual algal species of many shapes and sizes. There are about 130 genera in this group of unicellular microorganims, with about 2000 living and 2000 fossil species described so far.
The name "dinoflagellate" refers to the forward- spiraling swimming motion of these organisms. They are free-swimming protists (unicellular eukaryotic microorganisms) with two flagella, a nucleus with condensed chromosomes, chloroplasts, mitochondria, and Golgi bodies. Biochemically, photosynthetic species possess green pigments, chlorophylls a and c, and golden brown pigments, including peridinin. Dinoflagellates primarily exhibit asexual cell division, some species reproduce sexually, while others have unusual life cycles. Their nutrition varies from autotrophy (photosynthesis; in-nearly 50% of the known species) to heterotrophy (absorption of organic matter) to mixotrophy (autotrophic cells engulf other organisms, including other dinoflagellates).
Free-living dinoflagellates are an ancient and successful group of aquatic organisms. They have adapted to pelagic (free-floating) and benthic (attached) habitats from arctic to tropical seas, and to salinities ranging from freshwater, to estuaries, to hypersaline waters. Many species are found in numerous habitats, living in the plankton or attached to sediments, sand, corals, or to macroalgal surfaces or to other aquatic plants. Some species are present as parasites in marine invertebrates and fish. Some even serve as symbionts, known as zooxanthellae, providing organic carbon to their hosts: reef-building corals, sponges, clams, jellyfish, anemones and squid.
Dinoflagellates exhibit a wide variety in morphology and size (from 0.01 to 2.0 mm). They commonly have a cell covering structure (theca) that differentiates them from other algal groups. Cells are either armored or unarmored. Armored species have thecae divided into plates composed of cellulose or polysaccharides which are key features used in their identification. The cell covering of unarmored species is comprised of a membrane complex. The theca can be smooth and simple or laced with spines, pores and/or grooves and can be highly ornamented.
In systematics, dinoflagellates have been claimed by both botanists and zoologists. Dinoflagellates share features common to both plants and animals: they can swim, many have cell walls, and both photosynthetic and nonphotosynthetic species are known. Botanists have grouped them with the "microalgae" and zoologists have grouped them with the protozoa, and both have produced classification schemes for this diverse and confusing group.
Dinoflagellates have attracted a lot of negative attention from the general public in recent times. For example, blooms (population explosions) of dinoflagellates can cause the water to turn a reddish-brown color known as "red tide". Red tides can have harmful effects on the surrounding sea-life and their consumers. Additionally, certain species of dinoflagellates produce neurotoxins. These toxins are carried up the food chain, ultimately to humans and can, sometimes result in permanent neurological damage or even death. Yet dinoflagellates are important members of the phytoplankton in marine and freshwater ecosystems.
PHAEOPHYTA (brown algae)
The Phaeophyta are almost entirely marine, frequently dominating rocky shores in cold and temperate waters throughout the world. The giant kelp, Macrocystis pyrifera , forms expansive seaweed forests off the west coast of North America and provides habitat and shelter for many other organisms. Tropical waters have fewer species of brown algae, although genera such as Sargassum and Turbinaria can dominate in some areas to form small-scale forests. Sargassum is also unique among macroalgal genera in that it contains totally free-floating species with no requirement for attachment to the bottom, as in the Sargasso Sea.
The colors of brown algae (predominantly due to the brown accessory pigment fucoxanthin) cover a spectrum from pale beige to yellow-brown to almost black. In tropical seas, they range in size from microscopic filaments to several meters in length.
The large kelps are harvested for use as an emulsion stabilizer, in products such as ice cream. They are also used as fertilizer and as a vitamin rich food source
RHODOPHYTA (red algae)
Of the approximately 6000 species, most red algae are marine; only a few occur in freshwater. Rhodophytes are usually multicellular and grow attached to rocks or other algae, but there are some unicellular or colonial forms. They do not have flagellated cells, are structurally complex, and have complex life cycles divided into three phases. Many red algae feature pit connections between the cells, and their cell walls include a rigid component composed of microfibrils and a mucilaginous matrix. Agar and carrageenin are two red algal mucilages that are widely used for gelling and thickening purposes in the food and pharmaceutical industries.
Rhodophytes contain chlorophyll a which is masked by phycobilin pigments bound to proteins. The chloroplasts in red algae resemble Cyanobacteria both biochemically and structurally. Food reserves are stored outside of the chloroplasts as Floridean starch.
The coralline red algae deposit calcium carbonate in their cell walls, making them particularly tough and stony. They are often abundant, ecologically important, and widespread from the arctic regions to the tropics and play an important role in building tropical reef communities. Coralline red algae can form an algal ridge that absorbs wave energy and thereby protects the more delicate organisms that inhabit the sheltered lagoons and back-reef habitats.
Research scientist and curator Dr. Walter H. Adey has extensively collected the crustose corallines throughout the world since the 1960's, most notably in the North Atlantic, Caribbean and Hawaii. The specimens from the northwest Atlantic, north Norway and Iceland have been curated and inventoried during 2000-2001 with the assistance of Sue Lutz (Research and Curatorial Assistant). The specimens from Japanese regions, as well as the remaining European collections, are currently being curated and inventoried.
Some red algae are of economic importance, either as food (Porphyra) or as producers of secondary products (Gelidium, Gracilaria, Chondrus, etc.) used in the food and drug industries
As we have already read about bacteria , now we will focus deeply on algae so lets start studying about algae
what is algae ?
Algae are photosynthetic organisms that occur in most habitats. They vary from small, single-celled forms to complex multicellular forms, such as the giant kelps that grow to 65 meters in length.. The US Algal Collection is represented by almost 300,000 accessioned and inventoried herbarium specimens
Introduction Algae are photosynthetic organisms that occur in most habitats, ranging from marine and freshwater to desert sands and from hot boiling springs to snow and ice. They vary from small, single-celled forms to complex multicellular forms, such as the giant kelps of the eastern Pacific that grow to more than 60 meters in length and form dense marine forests. Algae are found in the fossil record dating back to approximately 3 billion years in the Precambrian. They exhibit a wide range of reproductive strategies, from simple, asexual cell division to complex forms of sexual reproduction.
Algae are important as primary producers of organic matter at the base of the food chain. They also provide oxygen for other aquatic life. Algae may contribute to mass mortality of other organisms, in cases of algal blooms, but they also contribute to economic well- being in the form of food, medicine and other products. In tropical regions, coralline algae can be as important as corals in the formation of reefs.
Seaweeds are larger algae that live in the marine (salt or brackish water) environment. Kelps are large brown seaweeds in the genera Pelagophycus, Laminaria, Macrocystis, etc. In the Pacific, individual kelp plants may reach 65 meters in length.
CLICK ON THE FIGURE BELOW FOR THE TOPICS IN WHICH YOU ARE INTERESTED
FASCINATING FACTS FROM BIOLOGYIf you yelled for 8 years, 7 months, and 6 days, you would have
produced enough sound energy to heat up one cup of coffee. (Hardly
seems worth it.)
A pig's orgasm lasts for 30 minutes.
(In my next life I want to be pig.)
Banging your head against a wall uses 150 calories an hour.
(Still not over that pig thing.)
Humans and dolphins are the only species that have sex for pleasure.
(Is that why Flipper is always smiling? And why isn't the pig
included in this list? Maybe 30 minute orgasms aren't as fun as I
imagine...)
On average, people fear spiders more then they do death.
The strongest muscle in the body is the tongue. (Hmmmmmm......)
A crocodile cannot stick out its tongue.
The ant can lift 50 times its own weight, can pull 30 times its own
weight, and always falls over on its right side when intoxicated.
(From drinking little bottles of....? Did the gov't pay for this
research?)
Polar bears are left handed.
(Who knew? Who cares? Did the gov't pay for this too? Probably...)
The flea can jump 350 times its body length. That's like a human
jumping the length of a football field.
A cockroach will live 9 days without its head before it starves to
death.
The male praying mantis cannot copulate while it's head is attached
to its body. The female initiates sex by ripping the male's head
off. (Hi, honey. I'm home. What the...?)
Some lions mate over 50 times a day.
(In my next life I still want to be a pig.
Quality over quantity, you know?)
Butterflies taste with their feet. (Oh, jeez!)
Elephants are the only animals that can't jump.
An ostrich's eye is bigger than its brain.
(I know some people like that.)
Starfish don't have brains.
(I know some people like this too.)
It takes your food seven seconds to get from your mouth to your stomach.
One human hair can support 6.6 pounds.
The average man's penis is two times the length of his thumb.
Human thighbones are stronger than concrete.
A woman's heart beats faster than a man's.
There are about one trillion bacteria on each of your feet.
Women blink twice as often as men.
The average person's skin weighs twice as much as the brain.
Your body uses 300 muscles to balance itself when you are standing still.
If saliva cannot dissolve something, you cannot taste it.
Women Will be finished reading this by now.
And Men Are still busy checking their thumbs.
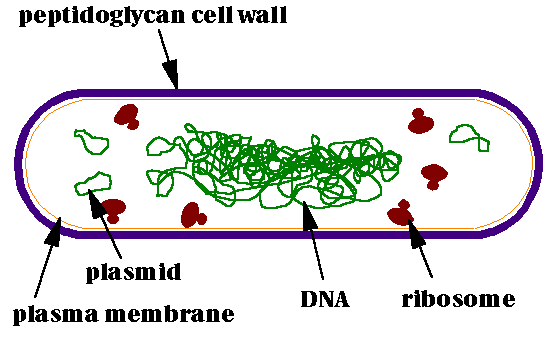
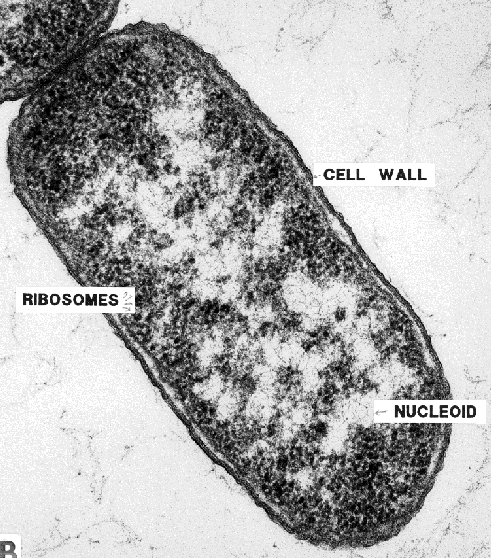
A more or less typical bacterium, shown here, is comparatively much simpler than a typical eukaryotic cell. View the transmission electron micrograph of a typical bacterium, E. coli, below and compare it with the diagram above.
Bacteria lack the membrane-bound nuclei of eukaryotes; their DNA forms a tangle known as a nucleoid, but there is no membrane around the nucleoid, and the DNA is not bound to proteins as it is in eukaryotes. Whereas eukaryote DNA is organized into linear pieces, the chromosomes, bacterial DNA forms loops. Bacteria contain plasmids, or small loops of DNA, that can be transmitted from one cell to another, either in the course of sex (yes, bacteria have sex) or by viruses. This ability to trade genes with all comers makes bacteria amazingly adaptible; beneficial genes, like those for antibiotic resistance, may be spread very rapidly through bacterial populations. It also makes bacteria favorites of molecular biologists and genetic engineers; new genes can be inserted into bacteria with ease.
Bacteria do not contain membrane-bound organelles such as mitochondria or chloroplasts, as eukaryotes do. However, photosynthetic bacteria, such as cyanobacteria, may be filled with tightly packed folds of their outer membrane. The effect of these membranes is to increase the potential surface area on which photosynthesis can take place.
The cell membrane is surrounded by a cell wall in all bacteria except one group, the Mollicutes, which includes pathogens such as the mycoplasmas. The composition of the cell wall varies among species and is an important character for identifying and classifying bacteria. In this diagram, the bacterium has a fairly thick cell wall made of peptidoglycan (carbohydrate polymers cross-linked by proteins); such bacteria retain a purple color when stained with a dye known as crystal violet, and are known as Gram-positive (after the Danish bacteriologist who developed this staining procedure). Other bacteria have double cell walls, with a thin inner wall of peptidoglycan and an outer wall of carbohydrates, proteins, and lipids. Such bacteria do not stain purple with crystal violet and are known as Gram-negative.
Classifying bacteria on the basis of their morphology is extremely difficult; bacteria are generally quite small and have simple shapes, though there are some bacteria, notably the cyanobacteria and actinomycetes, with sufficiently complex morphology to permit classification by shape. In addition to shape, bacteria have traditionally been identified and classified on the basis of their biochemistry and the conditions under which they grow. The advent of molecular biology has made it possible to classify bacteria on the basis of similarities among DNA sequences, and has revolutionized thinking in bacterial systematics. The cladogram above is based on DNA sequences that encode ribosome structure.
Bacteria grow in a wide variety of habitats and conditions.
When most people think of bacteria, they think of disease-causing organisms, like the Streptococcus bacteria growing in culture in this picture, which were isolated from a man with strep throat. While pathogenic bacteria are notorious for such diseases as cholera, tuberculosis, and gonorrhea, such disease-causing species are a comparatively tiny fraction of the bacteria as a whole.
Bacteria are so widespread that it is possible only to make the most general statements about their life history and ecology. They may be found on the tops of mountains, the bottom of the deepest oceans, in the guts of animals, and even in the frozen rocks and ice of Antarctica. One feature that has enabled them to spread so far, and last so long is their ability to go dormant for an extended period.
Bacteria have a wide range of envronmental and nutritive requirements.
Most bacteria may be placed into one of three groups based on their response to gaseous oxygen. Aerobic bacteria thrive in the presence of oxygen and require it for their continued growth and existence. Other bacteria are anaerobic, and cannot tolerate gaseous oxygen, such as those bacteria which live in deep underwater sediments, or those which cause bacterial food poisoning. The third group are the facultative anaerobes, which prefer growing in the presence of oxygen, but can continue to grow without it.
Bacteria may also be classified both by the mode by which they obtain their energy. Classified by the source of their energy, bacteria fall into two categories: heterotrophs and autotrophs. Heterotrophs derive energy from breaking down complex organic compounds that they must take in from the environment -- this includes saprobic bacteria found in decaying material, as well as those that rely on fermentation or respiration.
The other group, the autotrophs, fix carbon dioxide to make their own food source; this may be fueled by light energy (photoautotrophic), or by oxidation of nitrogen, sulfur, or other elements (chemoautotrophic). While chemoautotrophs are uncommon, photoautotrophs are common and quite diverse. They include the cyanobacteria, green sulfur bacteria, purple sulfur bacteria, and purple nonsulfur bacteria. The sulfur bacteria are particularly interesting, since they use hydrogen sulfide as hydrogen donor, instead of water like most other photosynthetic organisms, including cyanobacteria.
Bacteria play important roles in the global ecosystem.
The ecosystem, both on land and in the water, depends heavily upon the activity of bacteria. The cycling of nutrients such as carbon, nitrogen, and sulfur is completed by their ceaseless labor.
Organic carbon, in the form of dead and rotting organisms, would quickly deplete the carbon dioxide in the atmosphere if not for the activity of decomposers. This may not sound too bad to you, but realize that without carbon dioxide, there would be no photosynthesis in plants, and no food. When organisms die, the carbon contained in their tissues becomes unavailble for most other living things.Decomposition is the breakdown of these organisms, and the release of nutrients back into the environment, and is one of the most important roles of the bacteria.
The cycling of nitrogen is another important activity of bacteria. Plants rely on nitrogen from the soil for their health and growth, and cannot acquire it from the gaseous nitrogen in the atmosphere. The primary way in which nitrogen becomes available to them is through nitrogen fixation by bacteria such as Rhizobium, and by cyanobacteria such as Anabaena, Nostoc, and Spirulina, shown at right. These bacteria convert gaseous nitrogen into nitrates or nitrites as part of their metabolism, and the resulting products are released into the environment. Some plants, such as liverworts, cycads, and legumes have taken special advantage of this process by modifying their structure to house the basteria in their own tissues. Other denitrifying bacteria metabolize in the reverse direction, turning nitrates into nitrogen gas or nitrous oxide. When colonies of these bacteria occur on croplands, they may deplete the soil nutrients, and make it difficult for crops to grow.
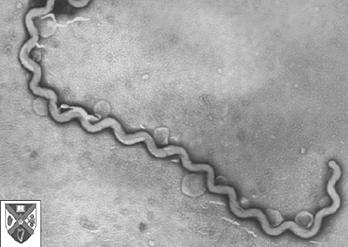
It may seem surprising that bacteria can leave fossils at all. However, one particular group of bacteria, the cyanobacteria or "blue-green algae," have left a fossil record that extends far back into the Precambrian - the oldest cyanobacteria-like fossils known are nearly 3.5 billion years old, among the oldest fossils currently known. Cyanobacteria are larger than most bacteria, and may secrete a thick cell wall. More importantly, cyanobacteria may form large layered structures, called stromatolites (if more or less dome-shaped) or oncolites (if round). These structures form as a mat of cyanobacteria grows in an aquatic environment, trapping sediment and sometimes secreting calcium carbonate. When sectioned very thinly, fossil stromatolites may be found to contain exquisitely preserved fossil cyanobacteria and algae.
The picture above is a short chain of cyanobacterial cells, from the Bitter Springs Chert of northern Australia (about 1 billion years old). Very similar cyanobacteria are alive today; in fact, most fossil cyanobacteria can almost be referred to living genera. Compare this fossil cyanobacterium with this picture of the living cyanobacterium Oscillatoria:
The group shows what is probably the most extreme conservatism of morphology of any organisms.
Aside from cyanobacteria, identifiable fossil bacteria are not particularly widespread. However, under certain chemical conditions, bacterial cells can be replaced with minerals, notably pyrite or siderite (iron carbonate), forming replicas of the once-living cells, or pseudomorphs. Some bacteria secrete iron-coated sheaths that sometimes fossilize. Others may bore into shells or rocks and form microscopic canals within the shell; such bacteria are referred to as endolithic, and their borings can be recognized all through the Phanerozoic. Bacteria have also been found in amber -- fossilized tree resin -- and in mummified tissues. It is also sometimes possible to infer the presence of disease-causing bacteria from fossil bones that show signs of having been infected when the animal was alive. Perhaps most amazing are the fossils left by magnetobacteria -- a group of bacteria which form tiny, nanometer-sized crystals of magnetite (iron oxide) inside their cells. Magnetite crystals identifiable as bacterial products have been found in rocks as old as two billion years -- at a size of a few hundred millionths of a meter, these hold the record for the smallest fossils.
Bacteria are often maligned as the causes of human and animal disease (like this one, Leptospira, which causes serious disease in livestock). However, certain bacteria, the actinomycetes, produce antibiotics such as streptomycin and nocardicin; others live symbiotically in the guts of animals (including humans) or elsewhere in their bodies, or on the roots of certain plants, converting nitrogen into a usable form. Bacteria put the tang in yogurt and the sour in sourdough bread; bacteria help to break down dead organic matter; bacteria make up the base of the food web in many environments. Bacteria are of such immense importance because of their extreme flexibility, capacity for rapid growth and reproduction, and great age - the oldest fossils known, nearly 3.5 billion years old, are fossils of bacteria-like organisms.
Click on the buttons below to learn more about the bacteria
That chameleons change colors to disguise themselves is a common misconception. Scientists believe that the real reason for color changing is to communicate with other chameleons and express their mood, like a reptilian mood ring! If the color happens to match the background, it's completely coincidental.
The also speculate that another changing colors could be advantageous is that it lets them adjust to various temperatures and light situations. For example, if the chameleon is cold, it will change to a darker color to absorb more light!
1. There are 62,000 miles of blood vessels in the human body – laid end to end they would circle the earth 2.5 times
2. At over 2000 kilometers long, The Great Barrier Reef is the largest living structure on Earth
3. The risk of being struck by a falling meteorite for a human is one occurrence every 9,300 years
4. A thimbleful of a neutron star would weigh over 100 million tons
5. A typical hurricane produces the energy equivalent of 8,000 one megaton bombs
6. Blood sucking hookworms inhabit 700 million people worldwide
7. The highest speed ever achieved on a bicycle is 166.94 mph, by Fred Rompelberg
8. We can produce laser light a million times brighter than sunshine
9. 65% of those with autism are left handed
10. The combined length of the roots of a Finnish pine tree is over 30 miles
11. The oceans contain enough salt to cover all the continents to a depth of nearly 500 feet
12. The interstellar gas cloud Sagittarius B contains a billion, billion, billion liters of alcohol [JFrater is planning to move there in the near future]
13. Polar Bears can run at 25 miles an hour and jump over 6 feet in the air
14. 60-65 million years ago dolphins and humans shared a common ancestor
15. Polar Bears are nearly undetectable by infrared cameras, due to their transparent fur
16. The average person accidentally eats 430 bugs each year of their life
17. A single rye plant can spread up to 400 miles of roots underground
18. The temperature on the surface of Mercury exceeds 430 degrees C during the day, and, at night, plummets to minus 180 degrees centigrade
19. The evaporation from a large oak or beech tree is from ten to twenty-five gallons in twenty-four hours
20. Butterflies taste with their hind feet, and their taste sensation works on touch – this allows them to determine whether a leaf is edible
The Difference Between Insects And Spiders ? Despite the visual similarities between the two both are actually members of distinct families. Spiders are members of the Arachnid family whilst insects belong to the Insect family. So what makes the two families different ? Well for starters spiders have 8 legs whilst insects have just 6. Insects have 3 distinct body parts, a head, thorax and abdomen whilst spiders have just 2, a combined head and thorax known as the cephalothorax and an abdomen. As well as the above, other interesting differences include the eyes. Spiders have simple eyes whilst insects have compound eyes. Spiders have piercing jaws whilst insects have jaws which are more suitable for chewing. All spiders can make silk thread (although not all spiders spin webs) whilst most insects cannot. Spiders can’t fly where as many insects have wings allowing them to do so. So yes to the untrained eye there is indeed very little visual difference but if you look really hard then they’re actually worlds apart.
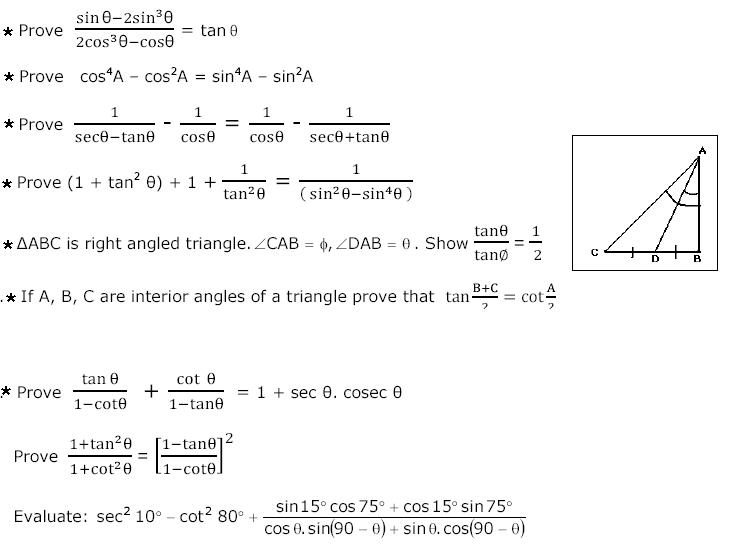
Now that we have polar basis vectors under our wing (and the polar representations of velocity and angular momentum), we are ready to proceed with the proof of Kepler's first law -- that the orbits of planets are ellipses with the Sun at one focus.To begin with, we will start off by applying Newton's law of motion and Newton's law of universal gravitation together to find that
ma = (-G m M/r2) r
equation 1
and, dividing both sides of the equation by m,
a = (-G M/r2) r.
equation 2
Recalling our work with polar basis vectors, we know dtheta/dtheta = -r. Solving for r and applying the chain rule, we find that
r = -dt/dtheta dtheta/dt
equation 3
r = -1/omega dtheta/dt.
equation 4
Substituting this into our equation for a we find
a = (-G M/r2) (-1/omega) dtheta/dt
equation 5
a = (G M)/(r2omega) dtheta/dt.
equation 6
If we multiply the right side of the equation by m/m (which is unity), we obtain
a = (G m M)/(m r2omega) dtheta/dt.
equation 7
But l = m r2omega (I told you this would come in handy as well), so we can rewrite this as
a = (G m M/l) dtheta/dt.
equation 8
We can multiply both sides by l/(G m M) and find
l/(G m M) a = dtheta/dt.
equation 9
But we know that a = dv/dt, and can substitute accordingly:
l/(G m M) dv/dt = dtheta/dt.
equation 10
This is a differential equation that we can now solve. Upon solving it, we find that
l/(G m M) v = theta + C
equation 11
where C is some constant vector. We'll solve this for v and find that
v = (G m M/l) (theta + C).
equation 12
This is a general solution to the differential equation.But we're not finished. This doesn't tell us much about the shape of a planet's orbit, although all the pieces are there. This is the general solution, and it could be an orbit of any of the possible shapes (though we can't be sure what they are yet) or any of the possible orientations. We're interested in knowing the shape, of course, so we want to restrict the possible orientations. To do that, we'll take a special case. It makes sense to have perihelion -- that is, closest approach to the Sun -- at time t = 0. We'll restrict the orientation so that, when perihelion occurs, the planet lies along the zero radian line from the Sun (or, in Cartesian terminology, along the positive x-axis) -- that is, theta = 0. At this point, r, the position vector of the planet, will have only a component in the positive x-axis. We'll also assume that the planet orbits the Sun counterclockwise, through increasing measures of angles. If this is the case, then the velocity v at the instant of perihelion should be orthogonal to the position vector r, and it should have only a component in the positive y-axis. According to our expression for v, we have a scalar times the vector quantity theta + C.
theta|(theta = 0) = j;
equation 13
that is, the unit transverse vector points "up" when the unit radial vector points "right." Since, at t = 0, theta points entirely in the y-direction, then our constant vector C must only have a component in the y-axis -- this is the only way to get a resultant vector (v) that points entirely in the y-direction. So, we can rewrite C as a scalar times the unit basis vector in the y-direction:
C = ej
equation 14
where e is some scalar constant. (You will find that we have a very good reason for choosing the letter e in this case.) Substituting this into our equation for v, we get
v = (G m M/l) (theta + ej).
equation 15
This is the specific case when we want the orbit oriented so that perihelion occurs at t = theta = 0.Now we are ready to finish up the problem. We can dot both sides of the equation with theta and get:
v dot theta = (G m M/l) (theta + ej) dot theta
equation 16
v dot theta = (G m M/l) [theta dot theta + e (j dot theta)].
equation 17
A vector dotted with itself yields the square of that vector's magnitude, so theta dot theta = 1. Simplifying v dot theta, we find
v dot theta = (dr/dtr + r omegatheta) dot theta
equation 18
v dot theta = dr/dt (r dot theta) + r omega (theta dot theta).
equation 19
But the dot product of two orthogonal vectors is zero, so r dot theta = 0. We also already know that theta dot theta = 1. Therefore
v dot theta = r omega.
equation 20
The last part of our problem is finding an expression for j dot theta. We know that theta = -sin theta i + cos thetaj (by definition), so
j dot theta = j dot (-sin theta i + cos thetaj)
equation 21
j dot theta = cos theta.
equation 22
We have the three pieces of the puzzle, so we'll put them together to find that
r omega = (G m M/l) (1 + e cos theta).
equation 23
We see r omega on the left side of this equation. We know that l = mr2omega, and it would be nice to get rid of the mess that way. So we'll multiply both sides of the equation by m r to get
mr2omega = (Gm2M/l) [r (1 + e cos theta)].
equation 24
Replacing the left side of the equation by l and moving the constants to the left side of our equation, we find that
l = (Gm2M/l) [r (1 + e cos theta)].
equation 25
We're almost finished now. Here it is clear that we have an explicit function in terms of r and theta -- in other words, this should be the polar equation for our planet's orbit! All we need do now is solve for r:
r = [l2/(Gm2M)]/(1 + e cos theta).
equation 26
The equation of a conic section with focus-directrix distance p and eccentricity e is represented by the polar equationa
r = e p/(1 + e cos theta).
equation 27
But this is exactly what we have, given that
e p = l2/(Gm2M).
equation 28
The focus-directrix distance should be a constant, which p is: l, G, m, and M are all individually constant; therefore the expression l2/(Gm2M) must also be constant. Therefore, Newton's laws of motion and universal gravitation dictate that the orbits of planets follow conic sections. This is Kepler's first law.... Well, almost. Kepler's first law actually states that planets follow the paths of ellipses. An ellipse is only one type of conic section. So why is an ellipse allowed while the others are not? Because the others are allowed -- we just don't call them planets. When Kepler said planet, he meant a body which returns to our skies over and over again. This means that the curve representing the orbit must be closed -- that is, it must continue to retrace itself over and over. The only two conic sections which are closed are the circle and the ellipse (the circle is just a special case of the ellipse anyway). Thus, any body in the skies we see over and over must be orbiting the Sun in an ellipse. The other two conic sections -- the parabola and hyperbola -- are open curves and correspond to a position where the body has sufficient velocity to escape from the Sun's gravity well. The body would approach the Sun from an infinite distance, round the Sun rapidly, and then recede away into the infinite abyss, never to be seen again. So we have proved an extension of Kepler's first law: A body influenced by the Sun's gravity follows a path defined by a conic section with the Sun at one focus.






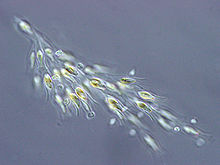
 Even though the Cyanobacteria are classified as bacteria (lacking a membrane-bounded nucleus) they are photosynthetic and are included among our algal collections. Cyanobacteria played a decisive role in elevating the level of free oxygen in the atmosphere of the early Earth. Cyanobacteria can change remarkably in appearance, depending on the environmental conditions. Blue-green algae are common in soil, in both salt and fresh water, and can grow over a wide range of temperatures. They have been found to form mats in Antarctic lakes under several meters of ice and are responsible for the beautiful colors of the hot springs at Yellowstone and elsewhere. Cyanobacteria can also occur as symbionts of protozoans, diatoms and lichen-forming fungi, and vascular plants. Some blue-greens can fix nitrogen as well as photosynthesize, allowing them to grow with only light, water, a few minerals, and the nitrogen and carbon dioxide in the atmosphere.
Even though the Cyanobacteria are classified as bacteria (lacking a membrane-bounded nucleus) they are photosynthetic and are included among our algal collections. Cyanobacteria played a decisive role in elevating the level of free oxygen in the atmosphere of the early Earth. Cyanobacteria can change remarkably in appearance, depending on the environmental conditions. Blue-green algae are common in soil, in both salt and fresh water, and can grow over a wide range of temperatures. They have been found to form mats in Antarctic lakes under several meters of ice and are responsible for the beautiful colors of the hot springs at Yellowstone and elsewhere. Cyanobacteria can also occur as symbionts of protozoans, diatoms and lichen-forming fungi, and vascular plants. Some blue-greens can fix nitrogen as well as photosynthesize, allowing them to grow with only light, water, a few minerals, and the nitrogen and carbon dioxide in the atmosphere.  Cyanobacteria are different in many important ways from other photosynthetic prokaryotes. Instead of the bacteriochlorophylls found in purple and green bacteria, blue-greens contain chlorophyll a, as in eucaryotic phototrophs, and, produce free oxygen as a byproduct of photosynthesis. Cyanobacteria, however, lack the organized chloroplasts of eukaryotes and have their photosynthetic apparatus distributed peripherally in the cytoplasm.
Cyanobacteria are different in many important ways from other photosynthetic prokaryotes. Instead of the bacteriochlorophylls found in purple and green bacteria, blue-greens contain chlorophyll a, as in eucaryotic phototrophs, and, produce free oxygen as a byproduct of photosynthesis. Cyanobacteria, however, lack the organized chloroplasts of eukaryotes and have their photosynthetic apparatus distributed peripherally in the cytoplasm.




 If you yelled for 8 years, 7 months, and 6 days, you would have
produced enough sound energy to heat up one cup of coffee. (Hardly
seems worth it.)
A pig's orgasm lasts for 30 minutes.
(In my next life I want to be pig.)
Banging your head against a wall uses 150 calories an hour.
(Still not over that pig thing.)
Humans and dolphins are the only species that have sex for pleasure.
(Is that why Flipper is always smiling? And why isn't the pig
included in this list? Maybe 30 minute orgasms aren't as fun as I
imagine...)
On average, people fear spiders more then they do death.
The strongest muscle in the body is the tongue. (Hmmmmmm......)
A crocodile cannot stick out its tongue.
The ant can lift 50 times its own weight, can pull 30 times its own
weight, and always falls over on its right side when intoxicated.
(From drinking little bottles of....? Did the gov't pay for this
research?)
Polar bears are left handed.
(Who knew? Who cares? Did the gov't pay for this too? Probably...)
The flea can jump 350 times its body length. That's like a human
jumping the length of a football field.
A cockroach will live 9 days without its head before it starves to
death.
The male praying mantis cannot copulate while it's head is attached
to its body. The female initiates sex by ripping the male's head
off. (Hi, honey. I'm home. What the...?)
Some lions mate over 50 times a day.
(In my next life I still want to be a pig.
Quality over quantity, you know?)
Butterflies taste with their feet. (Oh, jeez!)
Elephants are the only animals that can't jump.
An ostrich's eye is bigger than its brain.
(I know some people like that.)
Starfish don't have brains.
(I know some people like this too.)
It takes your food seven seconds to get from your mouth to your stomach.
One human hair can support 6.6 pounds.
The average man's penis is two times the length of his thumb.
Human thighbones are stronger than concrete.
A woman's heart beats faster than a man's.
There are about one trillion bacteria on each of your feet.
Women blink twice as often as men.
The average person's skin weighs twice as much as the brain.
Your body uses 300 muscles to balance itself when you are standing still.
If saliva cannot dissolve something, you cannot taste it.
Women Will be finished reading this by now.
And Men Are still busy checking their thumbs.
If you yelled for 8 years, 7 months, and 6 days, you would have
produced enough sound energy to heat up one cup of coffee. (Hardly
seems worth it.)
A pig's orgasm lasts for 30 minutes.
(In my next life I want to be pig.)
Banging your head against a wall uses 150 calories an hour.
(Still not over that pig thing.)
Humans and dolphins are the only species that have sex for pleasure.
(Is that why Flipper is always smiling? And why isn't the pig
included in this list? Maybe 30 minute orgasms aren't as fun as I
imagine...)
On average, people fear spiders more then they do death.
The strongest muscle in the body is the tongue. (Hmmmmmm......)
A crocodile cannot stick out its tongue.
The ant can lift 50 times its own weight, can pull 30 times its own
weight, and always falls over on its right side when intoxicated.
(From drinking little bottles of....? Did the gov't pay for this
research?)
Polar bears are left handed.
(Who knew? Who cares? Did the gov't pay for this too? Probably...)
The flea can jump 350 times its body length. That's like a human
jumping the length of a football field.
A cockroach will live 9 days without its head before it starves to
death.
The male praying mantis cannot copulate while it's head is attached
to its body. The female initiates sex by ripping the male's head
off. (Hi, honey. I'm home. What the...?)
Some lions mate over 50 times a day.
(In my next life I still want to be a pig.
Quality over quantity, you know?)
Butterflies taste with their feet. (Oh, jeez!)
Elephants are the only animals that can't jump.
An ostrich's eye is bigger than its brain.
(I know some people like that.)
Starfish don't have brains.
(I know some people like this too.)
It takes your food seven seconds to get from your mouth to your stomach.
One human hair can support 6.6 pounds.
The average man's penis is two times the length of his thumb.
Human thighbones are stronger than concrete.
A woman's heart beats faster than a man's.
There are about one trillion bacteria on each of your feet.
Women blink twice as often as men.
The average person's skin weighs twice as much as the brain.
Your body uses 300 muscles to balance itself when you are standing still.
If saliva cannot dissolve something, you cannot taste it.
Women Will be finished reading this by now.
And Men Are still busy checking their thumbs.









 That chameleons change colors to disguise themselves is a common misconception. Scientists believe that the real reason for color changing is to communicate with other chameleons and express their mood, like a reptilian mood ring! If the color happens to match the background, it's completely coincidental.
That chameleons change colors to disguise themselves is a common misconception. Scientists believe that the real reason for color changing is to communicate with other chameleons and express their mood, like a reptilian mood ring! If the color happens to match the background, it's completely coincidental.



